
Specimen description :
- Full specimen.
- Prone on matrix.
- Total size : 85 mm
- Lower Ordovician, Arenigian.
- Kunda horizon.
- Wolchow River, St Petersburg.
- Russia.
Dorsal view of the positive part.
Asaphus raniceps
This is a trilobite occuring in the Lower Ordovician, Arenigian, of the North-West of Russia. This is one of the 35+ species of the Asaphus genus which nicest specimens are collected and prepared by and at the Paleolab of St Petersburg. Historically, most of the Asaphids have been found and described in Northern Europe, but the exceptional preservation of the russian specimens lead many collectors to renew their former specimens.
Asaphus raniceps can be found in North-West of Russia (Kunda horizon), in Sweden and Estonia. In general, Asaphus raniceps is very similar to A. lamanskii from which the diagnose criteria derives.
|
Specimen description :
|
|
Dorsal view of the positive part.
|
Diagnosis :
|
Order
|
Asaphida
|
SALTER 1864
|
Order including 1/5 of all the species of Trilobites,
it mostly regroup librostoms of various morphology, in which the most advanced
families do present a ventral median suture early visible in the ontogeny
( "asaphoïd" protaspid larvæ).
Occurrence : Middle-Upper Cambrian boundary to upper Ordovician-lower Silurian.
|
|
Superfamily |
Asaphoidea |
BURMEISTER 1843 |
Occurrence: Middle Cambrian to Ordovician.
|
|
Family
|
Asaphidæ |
BURMEISTER 1843 |
Family tends toward loss of apparent segmentation of cephalon and pygidium, obsolescence of axial furrows and deep notching of posterior margin of hypostoma.
|
|
Subfamily |
Asaphinæ |
BURMEISTER 1843 |
|
|
Genus |
Asaphus |
BRONGNIART 1822 |
Average sized trilobites. Occurence : lower and middle Ordovician, Europe and Asia.
|
|
Species |
raniceps |
DALMAN (?)
|
Average and large sized Asaphus (max : 95 mm). Occurence : Lower Ordovician, Arenigian. Body oval, rather broad.
|
Notes :
When I published this page on the web, Serge Terentiev pointed out on the Yahoo trilobite 2 club the fact that the russian form of A. raniceps has been reviewed in recent publications "Asaphus ex.gr raniceps". He precised that better understanding of this problem could be read in a work of Arne T. Nielsen, 1995, "Trilobites of Hyk Formation and Komstad Limestone, Southern Sweden".
Dicrano then answered : "This is because a number of publications have questioned whether the Swedish and Russian "Asaphus raniceps" are the same species. There is a paper this year that addresses this issue. "The Asaphus raniceps problem in the Ordovician of Sweden." An abstract of it can be found here (NB : copied further down in case of breaking of the link). I have not seen the full paper yet."
I contacted Mr J. Bergström who gracefully sent me a reprint of his great work and for what I heartfully thank him. Here is a summary and the conclusion of this paper :
- The initial trouble comes from a drawing Angelin made from A. raniceps in 1854 (in a paper where he introduced A. fallax, bringing the number of Östergotland asaphids species to 3 in association with A. expansus WAHLENBERG 1818 and A. expansus var.ß raniceps DALMAN 1827, later renammed A. raniceps). Jaanusson in 1953 compared one of the drawing of Angelin to Dalman's concept of A. raniceps and concluded that this specimen did not fit the scheme. He was followed in this idea by Tjernvik & Jaanusson (1980), Nielsen (1995) et Bruton et al. (1997).
- Bergström, Zhi-Qiang, Zhi-Yi et Wen-Wei have rewiewed the collection studied by Tjernvik & Jaanusson (the large Holger Pihl collection) and concluded that there is no distinction between A. raniceps "in the Tjernvik & Jaanusson sense " (and not Angelin's one ;) ) and A. fallax. I report here 2 interesting systematic points (amongst other) that may have lead them in mistake (copy of the text) :
- "The deeply impressed glabellar muscle scars, thought by Nielsen (1995) to be typical of A. fallax are not so impressive in the lectotype (Nielsen 1995, fig. 62B) and are seen occasionnally in specimens from Ljungsbro, and occur in individuals of other Asaphus species as well. For instance, they are seen in some specimens of A. raniceps and they are strongly developped in the lectotype of A striatus (cf. Størmer 1940, pl.3, fig.12). Obviously it is not a reliable species character."
- " There is also individual variation in the development of the posterior border furrow. It can have a almost V-shaped cross section, but can also have a more rounded bottom as in A. expansus.".
- Finally, correcting a few other points concerning the drawings of Tjernvik & Jaanusson and Nielsen and a "pseudo-rarity" argument, J. Bergström et al. could conclude that "Angelin was correct in identifying Wahlenberg's and Dalman's species of Asaphus".[...] The situation can be summarised as follow :"
|
Name in current use
|
Correct Name
|
| A. expansus WAHLENBERG | A. expansus WAHLENBERG |
| A. fallax ANGELIN | A. fallax ANGELIN |
| A. raniceps sensu DALMAN | A. raniceps DALMAN |
| A. raniceps sensu ANGELIN | A. raniceps DALMAN |
Now, I'd like to once again heartfully thank Mr. J.Bergström for his help and say that his concise work is of great help in solving this systematic problem that occured with the asaphids of Sweden.
Here is the the abstract of the paper :
|
The Asaphus raniceps problem in the Ordovician of Sweden Bergström, J., Zhou Zhi-Qiang, Zhou Zhi-yi & Yuan Wen-Wei 20030430 : The Asaphus raniceps problem in the Ordovician of Sweden. Bulletin of Geological Society of Denmark, Vol. 50, pp. 95104, Copenhagen. It has been repeatedly stated that the lower Llanvirn Asaphus raniceps described by Dalman in 1827 and by Angelin in 1854 may represent two different species. This has forced us to live with the concepts of both A. raniceps Dalman and A. raniceps sensu Angelin, and with a zone of A. raniceps where A. raniceps is supposedly not present. However, new considerations based on large old and new collections have led to the firm conclusion that Angelin was correct in his identification of Dalmans species. What has been regarded recently as the true Asaphus raniceps, at least by Tjernvik & Johansson in 1980 and by Nielsen in 1995, is instead identical with another generally misidentified species, Asaphus fallax Angelin 1854. Key words: Ordovician, Baltoscandia, trilobites, Asaphus, Dalman, Angelin. Jan Bergström [jan.bergstrom@nrm.se], Swedish Museum of Natural History, P.O. Box 50007, SE-104 05 Stockholm, Sweden. Zhou Zhi-Quiang [xigmrx@pub.xaonline.com], Xian Institute of Geology and Mineral Resources, 166 East You Yi Road, Xian, Peoples Republic of China. Zhou Zhi-Yi [zyizhou@pub.jlonline.com] & Yuan Wen-Wei, Nanjing Institute of Geology and Palaeontology, Academia Sinica, 39 East Beijing Road, Nanjing 210008, Peoples Republic of China. 16 August 2001. |
Complementary pictures :
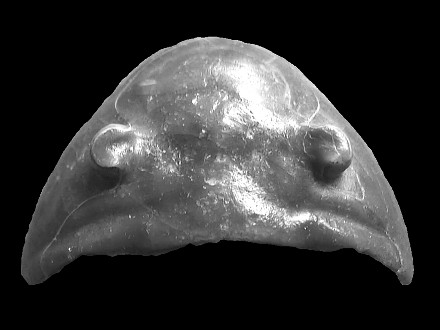 |
 |
|
Dorsal view of the cephalon
|
Dorsal view of the pygidium.
|
Well, so far for the "less expressed, discontinuous, segmentation" ... I have to admit my picture don't deserve well this point ;)
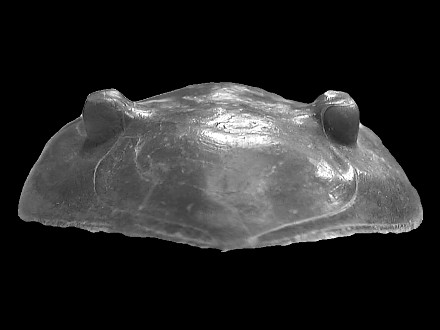 |
|
Frontal view of the cephalon. Note the lack of occipital
node.
|
 |
|
Side view of the complete specimen |